Коммуникация животных: от стимула к символу

Научное исследование коммуникации животных требует свободы от субъективизма и антропоморфизма в описании и в объяснении поведения. Исследователь не должен гадать, пусть даже убедительно, что именно животное хочет выразить данной демонстрацией, каковы его побуждения и т. д. Поэтому описание поведения, его разделение на элементы, проверку наличия у них сигнальной функции нужно производить с той же объективностью, с какой специалист по сравнительной анатомии выделяет морфоструктуры. Верно и обратное: только выделив устойчивые поведенческие паттерны, можно затем исследовать их возможные функции, в том числе и сигнальные. Прежде чем перейти к примерам, напомним основные понятия.
«Сигнальность» и «знаковость» демонстраций: история идеи
История исследования сигналов и механизмов коммуникации животных тесно связана с историей сравнительной этологии. В отличие от других поведенческих дисциплин (зоопсихологии, сравнительной психологии, физиологии высшей нервной деятельности), сравнительная этология, во-первых, анализирует именно видоспецифическое поведение, то есть самые жесткие, стереотипные, четко дифференцированные элементы последнего, во-вторых, кладет «морфологический» подход к поведению в основу своего метода.
Первыми здесь были учителя Конрада Лоренца — немецкий орнитолог, создатель этологического метода Оскар Хейнрот и английский биолог-эволюционист Джулиан Хаксли (Лоренц стажировался у него в 1920-е годы). Они попытались выделить в изменчивом поведении возбужденного животного (во время сексуальных или агрессивных взаимодействий) элементы, которые были бы столь же четки, дискретны и характерны для вида, как морфоструктуры. Лоренц легко воспринял эти идеи; недаром его первым научным руководителем был Фердинанд Хохштеттер, специалист по сравнительной и систематической морфологии, а также сравнительной эмбриологии позвоночных.
Оскар Хейнрот ввел представление о значимых элементах поведения — демонстрациях и ритуалах как специфических структурах, выделяемых в потоке деятельности животных, существенных для участников взаимодействия и допускающих «морфологическое» либо функциональное описание в той же степени, что и «обычные» морфоструктуры. Это был полный разрыв этологии с традиционным пониманием поведения животных как внешнего проявления «привычек», «инстинктов», «душевной деятельности» и иных эвфемизмов для обозначения психического. Он позволил этологам изучать сигнальные системы разных видов животных так же объективно, как лингвисты изучают языки. От традиционного же «психологического» понимания поведения (которое сохранили сравнительная психология, зоопсихология и физиология высшей нервной деятельности), этологи стремились всячески отмежеваться.
В 1911 году Хейнрот определил этологию как изучение «языка и ритуалов» животных, объединенных им в понятии «система коммуникации». Затем Джулиан Хаксли ввел понятие ритуализации, показав, что социальное поведение животных координируется с помощью сигналов, символизирующих определенную форму поведения, которая должна последовать за данной демонстрацией. Иначе говоря, демонстрация побуждений и намерений животного превращается в знак, указывающий на возможности развития поведения обоих участников.
Для Хаксли, Хейнрота и их общего ученика Лоренца это означало «презумпцию сигнальности» демонстраций. Всякое выразительное движение животного (то есть «демонстрации» в обыденном смысле слова) этологи рассматривали как потенциальный сигнал. Дальше следует естественное предположение, что «сигнальность» прямо пропорциональна стереотипности, фиксированное и демонстративности действий. Иначе зачем животному так коверкать и вычурно исполнять движения гнездостроения, агрессии, кормления, бегства и т. п., как не для того, чтобы подать сигнал партнеру? (Закрепление «коверканного», «вычурного», слишком шаблонного исполнения действий, происходящих из сферы повседневной активности, их необычное комбинирование друг с другом в комплексы как раз и составляет суть процесса ритуализации, превращающего действие в демонстрацию. Дальше мы рассмотрим примеры таких комплексов движений.)
Когда требуется не сигнал, а реакция — нападение или бегство, спаривание или оборона, прямые действия много эффективнее демонстраций. Последние из-за своей «неестественности» могут показывать только готовность к определенному поведению, тем самым побуждая партнера сделать свой «ход» — выбрать из репертуара брачных или агрессивных демонстраций «ответную». Первый партнер, в свою очередь, ответит на этот ход своим, и таким образом взаимодействие будет продвигаться к определенному исходу — победе одной особи и поражению другой, успешной копуляции и т. п. (Важно, что демонстрации не могут понудить партнера к чему-либо напрямую, как понуждает удар, клевок или иное прямое действие.) Неадекватные и неспецифические ответы ведут к остановке и срыву взаимодействия.
В чем заключалась сущность этологического подхода к выделению элементов поведения, обладающих потенциальной сигнальностью? Представим себе человека, на которого оказывают давление, требуя согласиться с чем-то неприемлемым: он нервно барабанит пальцами по столу, а затем, приняв решение, складывает из пальцев символическую фигуру, обозначающую отказ. Предполагалось, что в поведении животных можно точно так же выделить непрерывную последовательность реакций, обозначающих только возбуждение (аналог постукивания пальцами) и особые дискретные комплексы движений — демонстрации, которые, будучи продуктом ритуализации, по Хаксли, символизируют намерения участников «диалога». Их реакция на эти «символы» увеличивает предсказуемость поведения каждого и взаимно координирует поведение обоих. А это и есть цель коммуникации.
Важную роль в развитии этого направления сыграла работа Хейнрота, посвященная демонстрациям у разных видов утиных: выяснилось, что они строго видоспецифичны и имеют столь четко определенные формы, что наблюдатель (а значит, и другая птица того же вида) узнает каждую из них столь же безошибочно, как анатом — морфоструктуры. Затем Лоренц в посвященной Хейнроту работе 1941 года «Сравнительное исследование демонстраций утиных» показал, что сходство форм демонстраций отражает филогенетическую близость видов, и наоборот, чем более родственны виды друг другу, тем больше в их видовых репертуарах гомологичных демонстраций, различающихся лишь отдельными «оттенками» исполнения. Далее, у близких видов подавляющее большинство демонстраций с одним и тем же «смыслом» гомологичны или вообще тождественны по форме. Это позволяет говорить о том, что дивергенция (расхождение) близких видов одного рода или семейства сопровождается дивергенцией «словарей» видовых демонстраций. У неродственных видов репертуары демонстраций представлены в основном негомологичными сигналами.
Однако незначительные (с точки зрения наблюдателя!) различия между демонстрациями остаются устойчивыми, независимыми от колебаний состояния животного, если они важны для правильного восприятия другим животным. Это относится и к разным демонстрациям одного вида, и к гомологичным демонстрациям у разных видов.
Такие характерные отличия можно наблюдать у западной поганки Aechomorphus occidentalis (рис. 1). Демонстрации «dip-shaking» и «bob-shaking» обе используются во время ухаживания, их выразительные движения весьма сходны. Отличаются только порядок элементов и некоторые движения в ключевых моментах — в начале и конце демонстрации. «Dip-shaking» включает плескание водой в начале и позу с низкой шеей в конце, «bob-shaking» — нет. Это и делает их семантически различными сигналами: они обозначают разные степени преодоления взаимной агрессивности партнеров (а она неизменно присутствует в начале ухаживания, так же как и стремление к бегству) и разные возможности сближения.
Рис. 1. Демонстрации ухаживания «dip-shaking» и «bob-shaking» у водоплавающей птицы западной поганки очень сходны, но имеют отличия в ключевых моментах, в начале и конце. Этого достаточно, чтобы их визуальные образы четко различались. (Из: Nuechterlein С, Storer R., «Condor», 1982, т. 84. 4). Изображение: «Химия и жизнь»

Отметим, что птицы могут исполнять обе эти демонстрации также вне контекста образования пары — при агрессии и в других обстоятельствах, связанных с ростом возбуждения. Но тогда все «молекулы поведения» намного менее устойчивы. По мере подъема возбуждения особи видна сеть переходов между действиями повседневной активности и вычурными, экстравагантными движениями демонстраций, при спаде возбуждения — обратные переходы. Когда же демонстрации используются в специфическом контексте ухаживания, то есть как коммуникативные сигналы, — переходы не видны совсем. Стереотипное исполнение каждой демонстрации организовано таким образом, что одна сменяет другую скачком, как кадры слайд-фильма, и переходов не наблюдается, оно «подчеркивает» границы, отделяющие формы демонстраций друг от друга и от «фона» — несигнальной активности.
Рис. 2. Долгий крик у видов-двойников антарктических поморников: южнополярного ... Изображение: «Химия и жизнь»

Рис. 3. ... и бурого поморника (а—д — разные птицы, прорисовки по кадрам фильмов). Индивидуальная изменчивость не уничтожает общевидового облика демонстрации. Изображение: «Химия и жизнь»

Межвидовые различия демонстраций показаны у двух видов-двойников антарктических поморников — южнополярного поморника Catharacta maccormicki и бурого поморника C.lohnbergi (рис. 2, 3, 4) — на примере «долгого крика». Эта демонстрация многофункциональна, как, например, песня у воробьиных птиц. Для особей своего пола, с которыми конкурируют за партнера, она означает угрозу и стремление доминировать, для потенциального партнера — готовность поухаживать, для «нейтральных» особей — это сигнал, сплачивающий чаек в «клубах» и других группах. «Долгие крики» у обоих видов сходны, но между ними есть и четкие, устойчивые различия, как в целом, так и на каждом из пяти этапов: южнополярный поморник отклоняет корпус гораздо дальше назад и (или) вверх. Межиндивидуальная изменчивость «долгих криков» у обоих видов достаточно велика, но никогда не затрагивает видоспецифичных признаков. С другой стороны, стереотипность исполнения не ликвидирует и даже не уменьшает индивидуальную и ситуативную изменчивость — такое просто невозможно. Главное, что не изменяются дифференцирующие характеристики сигналов — те особенности, по которым различаются сигналы близких видов, как в примере с поморниками, или формы разных демонстраций ряда, как у поганок на рис. 1.
Рис. 4. Между «долгими криками» двух видов поморников сохраняются устойчивые различия: 1–5 — последовательные фазы долгого крика, столбики внизу — частота встречаемости у того и другого вида. (По:. Pietz P., «Condor», 1985, т. 81). Изображение: «Химия и жизнь»

Дискретно и дифференцированно
В примере со складыванием кукиша наличие переходов между элементами сигнального ряда не препятствует выделению отдельных демонстраций. Точно так же непрерывность мотивационных состояний не только не препятствует дискретности элементов демонстраций, но даже требует ее, если те функционируют как сигналы. В примерах, рассмотренных в предыдущей главе, половое возбуждение у демонстрирующей птицы нарастает плавно, однако демонстрации дискретны.
Вообще для коммуникации, то есть обмена сообщениями, несущими информацию, необходимо расчленение внешнего мира средствами некоторой семиотической системы (см., например: А. Н. Барулин, «Основы семиотики», 2002). Так, погода меняется непрерывно и плавно, но мы выделяем в этих изменениях значимые стадии и подбираем для них обозначения — «ясно», «переменная облачность», «дождь», будь то слова или пиктограммы в прогнозе погоды, изображающие солнце, тучку и т. п. (рис. 5). На основании такого сигнала другой человек может принять решение о том, как ему одеться перед выходом из дома. (Очевидно, что семиотическая система должна быть общей для приемников и передатчиков сигнала.)
Рис. 5. «Расчленение мира» для задач коммуникации. Погода изменяется плавно и непрерывно (а). Концептуализация — расчленение этого континуума на элементы: выделены значимые стадии изменений погоды, которые требуют по-разному одеваться (б). Именование (в) — обозначение элементов средствами семиотической системы (пиктограммами). Изображение: «Химия и жизнь»
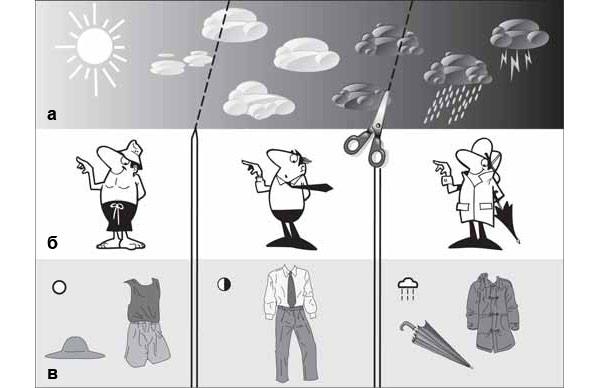
Если вернуться к коммуникации животных, то плавно меняющиеся мотивации можно сравнить с плавным изменением погоды, а демонстрации — с введением «обозначений», информирующих о состоянии животного. Континуум поведенческих характеристик животного или значимых деталей ситуации (предмет общения) дробится на конечное число дискретных уровней (означаемое), каждый из которых выражен собственным сигналом. Поведение особи-демонстратора должно разделиться на дискретные структуры — «сгустки» непрерывного потока активности, они же фиксированные комплексы действий, с дальнейшей ритуализацией каждого из них.
Так, у озерных чаек, которых исследовал Нико Тинберген, есть диагональная и распластанная позы угрозы, между которыми присутствуют всевозможные переходы. Киносъемка чаек показала, что в большинстве ситуаций нападения и бегства, во-первых, сами позы встречаются чаще, чем переходные движения, во-вторых, обе позы и переходы между ними — намного чаще, чем незавершенные попытки демонстрации поз. А главное, даже при наличии переходов позы отличаются вероятностями атаки и бегства оппонента после их предъявления — вторая влечет атаку с большей вероятностью. Иначе говоря, существуют два уровня реагирования оппонента, соответствующие первой и второй позе.
Другой пример: суслик Белдинга Spermophilus beldingi реагирует на потенциальную опасность тремя типами звуков — трелями, чириканьем и свистами. Трели, серии коротких, быстро следующих друг за другом звуков, суслик чаще всего издает в ответ на появление наземных хищников (в экспериментах — человека, собаки, чучел длиннохвостой ласки и американской норки). Одиночные чириканья и свисты чаще сопровождают появление пернатых хищников. Однако более детальный анализ показывает, что свистом обозначается всякое резкое движение, сильно пугающее животное, трелью — то, что пугает не очень сильно, неподвижная или, во всяком случае, контролируемая опасность. Чириканье же — переходный сигнал (1984. работы Д. Легера и соавторов).
Крайне существенно, что эти три типа сигналов, маркирующие три разные уровня беспокойства животного (о которых суслики «считают нужным сообщить» другим особям в группировке), устойчиво отделены друг от друга только тогда, когда они предупреждают об опасности. Однако тот же самый трелевой сигнал издают самцы сусликов после копуляции. Непосредственная причина этого — обычный для ухаживания конфликт между сексуальностью, страхом перед самкой и агрессией по отношению к ней, когда непосредственно перед копуляцией и во время нее уровень страха максимален. Но трели после копуляции употребляются с совершенно иным «синтаксисом» — они статистически достоверно отличаются от тех, что сигнализируют об опасности, меньшим числом элементов и большей длительностью каждого элемента, причем между двумя типами сигналов практически нет переходов. Кроме того, трели неспецифически маркируют высокое возбуждение животных — но в этом случае изменчивость сигнала настолько широка, что перекрывает оба «чистых» типа трелевого крика, и предупреждающий, и посткопуляторный, причем преобладают переходные формы.
Таким образом, мы убедились, что эффективная коммуникация требует специфических элементов поведения. Даже если в исходном поведении животного соответствующие поведенческие паттерны не разделены, то в контексте общения их приходится разделять.
Примеров, подтверждающих это заключение, сегодня известно гораздо больше, чем можно перечислить в популярной статье. Исследование демонстраций в рамках заложенной Хаксли идеи «сигнальности» и «знаковости» демонстраций оказалось весьма плодотворным (как писал продолживший их традиции Лоренц в «Оборотной стороне зеркала»). С одной стороны, были показаны дискретность и дифференцированность демонстраций — животное устойчиво воспроизводит определенные формы, как будто огоньком сигареты чертит фигуры в темноте, и эти «фигуры» демонстраций четко выделяются на фоне континуума несигнальных движений. Так, демонстрации угрозы у десятков исследованных видов птиц оказались дискретными и дифференцированными — градуальных сигналов не обнаружено. (Это было показано в обзоре П. Харда и М. Энгвиста по демонстрациям ритуализированной угрозы у птиц в Canadian J. Zool. (2001).) С другой стороны, установлено, что формы демонстраций, которые животное воспроизводит устойчиво и стереотипно, в самом деле имеют коммуникативное значение — именно на них ориентируется другое животное, корректируя свое поведение, а не, допустим, на изменения дистанции между особями, взаимной ориентации, интенсивности проявлений неритуализованной агрессии и т. п.
Рассказать подробно о каждом из этих исследований нет возможности, отметим лишь один важный момент. Коммуникация считалась доказанной, если реципиент изменял свое поведение под воздействием сигналов партнера при конкуренции за ресурс малой ценности. Например, в работах Энквиста и соавторов приведены результаты наблюдений за глупышами Fulmarus glacialis, оспаривающими потроха пинагора у мест, где выбрасывают рыбные отходы. Если ресурс не слишком важен, у животного есть выбор: продолжать борьбу или отказаться. Таким образом, этот ресурс делается пробным камнем, позволяющим выявить социально обусловленную разнокачественность особей. Это показывает, что истинная ценность коммуникации — не в ресурсе как таковом, а в выстраивании оптимальных отношений с компаньонами.
Знак и стимул в коммуникации животных
Возникает естественный вопрос: а как передается коммуникативный эффект? И тут есть две возможности. Первая — передача через обмен воздействиями. В этом случае сигналы будут стимулами, которые в классической этологии называют специфическими или знаковыми. Действуя на реципиента, они «принуждают» его к той ответной реакции, которая необходима на следующем этапе, реакция, в свою очередь, побуждает первого партнера к повышению стимуляции и т. д. — в результате действия партнеров становятся скоординированными. Подобное использование стимулов описывается в диалоговой модели коммуникации классических этологов, получившей название «модели ключа и замка». Стимул в этой модели называется релизером — калька немецкого Ausloesung: «высвобождение, срабатывание». Действие релизера высвобождает адекватный и специфический двигательный ответ реципиента, который тот выдает непроизвольно, автоматически, даже если он в это время занят чем-то другим, скажем, предъявляет свою демонстрацию. В другом варианте релизер может высвобождать несколько разных (альтернативных) ответов, каждый со своей вероятностью. Например, уже упоминавшиеся демонстрации озерных чаек — позы горизонтальная и распластанная, — обе вызывают как атаку, так и бегство партнера, но атака в ответ на вторую встречается чаще. Так или иначе, свобода выбора реагирования у обоих особей, и демонстрирующей, и воспринимающей, предельно ограничена, они могут или специфически отвечать на релизер, или «сопротивляться» ему, нарушая соответствие между стимулом и реакцией.
Если же набор демонстраций ухаживания, угрозы, предупреждения об опасности и т. п. — это система знаков, а дифференцированные сигналы соответствуют набору ситуаций взаимодействия, то они действуют исключительно информацией. И эта информация позволяет выбрать ответный сигнал таким образом, чтобы события развивались в направлении, более выгодном для данной особи, чем для ее оппонента. В этом случае свобода выбора у особи максимальна. С одной стороны, реципиент использует полученную информацию для корректировки собственной модели поведения там и тогда, где это необходимо именно для него (не обязательно сейчас). С другой стороны, сигнал не зависит от состояния обеих особей — передающей и принимающей, и поэтому воспринимается независимо от уровней испуга, агрессии и т. п.
Когда сигнальный эффект воздействия демонстраций передается стимулами, он информирует об уровне мотивации реципиента. Поэтому такие сигналы в англоязычной литературе получили название motivational signals (сигналы намерений и состояния животного). Когда же сигнальный эффект передается знаками, используется название referential signals. Эти сигналы имеют внешнего референта (адресата) и передают информацию о важных ситуациях взаимодействия или объектах внешнего мира (виды корма, типы опасностей). Информация закодирована в дискретных, дифференцированных, специфических формах сигналов, значение которых не зависит от контекста (один и тот же сигнал всегда означает одно и то же). Это было продемонстрировано в исследованиях 90-х годов (P.Marler, Ch.S.Evans, J.Macedonia, D.BIumstein), благодаря которым у самых разных видов позвоночных были обнаружены функционально референтные сигналы (referential signals), они же категориальные сигналы, они же сигналы-символы в нашей классификации. Они как раз и представляют собой знаки (и символы), передающие информацию идеального характера.
В изящных экспериментах Кристофера Эванса с коллегами, выполненных в 1990-е годы, было показано, что подобный сигнал информирует именно о категориях внешней опасности (о типе пищи, о форме угрозы и т. д.), а не о субъективном уровне возбуждения особи. У домашних кур описаны три сигнала — «опасность с земли», «опасность с воздуха» и пищевой крик. Сигналы четко дифференцированы, «промежуточные» звуки не отмечаются даже у сильно возбужденных птиц. Это позволяет предположить, что все они передают информацию об определенных ситуациях внешнего мира, которую адекватно воспринимают другие куры. Чтобы подтвердить или опровергнуть это предположение, нужно было проверить реакцию особей, которые сами не могут видеть опасность или корм — то есть им приходится полагаться именно и только на сигналы.
Для этого были поставлены две серии опытов, в которых курам проигрывали записи изучаемых криков. В контрольной серии (половина всех повторностей, выбранная случайным образом) проигрывание пищевого крика или крика «опасность с земли» предваряли выдачей зерна, причем птице давали возможность кормиться. В экспериментальной серии те же крики проигрывали в том же порядке, но им не предшествовала выдача зерна, то есть птицы ничем не были мотивированы «правильно» реагировать на тот и другой крик. Реакцию на сигнал оценивали по среднему времени поискового поведения в секундах (вертикальная ось на рисунке), при котором птица характерным образом меняет позу, опуская голову к земле (рис. 6). Понятно, что возможность высматривать хищника и реагировать на его появление при этом ограниченна. Поисковое поведение после сигналов сравнивали с таким же поведением в период до сигналов и оценивали приращение либо уменьшение.
Рис. 6. Сигналы-символы в действии: информирование о ситуации у домашних кур (из работы К. Эванса с соавт., 2007). Черные столбики — изменения интенсивности поискового поведения после проигрывания пищевого крика, белые — после крика «опасность с земли». На фото — поза поиска, которую принимает курица в ответ на пищевой крик именно при отсутствии подкрепления (то есть когда ничто не побуждает искать корм, кроме этого крика). Изображение: «Химия и жизнь»

Некоторый уровень пищевой активности у курицы есть всегда. Однако пищевой крик усиливает ее у голодной особи — птица «доверяет» пищевому сигналу, что надо тщательней искать именно здесь, хотя это может быть опасно из-за потери бдительности. Сытые птицы не так охотно реагируют на пищевой крик (то есть здесь нет «автоматической» реакции, подобной той, что вызывают релизеры). Тревожные сигналы не влияли на пищевое поведение ни у сытых, ни у голодных кур. Таким образом, пищевой сигнал действительно передает конкретную информацию: «здесь может быть еда, ищите лучше». А чтобы исследовать реакцию птиц на сигналы тревоги, нужны другие параметры — интенсивность осматривания, затаивания и т. п.
Предшествующие исследования тех же авторов показали то же самое и в отношении обоих «тревожных криков». Так, у цыплят домашней курицы специфическую реакцию затаивания и бегства вызывает не только крик «опасности с воздуха», но и стилизованное изображение ястреба — силуэт с широкими крыльями, короткой шеей и длинным хвостом, который двигают над цыплятами. Реакция на такой стимул была исследована еще в классических экспериментах Лоренца—Тинбергена. Эванс и соавторы повторили ее (1993), используя компьютерную анимацию движения «хищника» с разной скоростью и на разной высоте. В обоих случаях оборонительная реакция имела место, когда модель напоминала ястреба, и отсутствовала, когда ее двигали в противоположную сторону (тогда она напоминает утку — длинный «хвост» может быть интерпретирован как «шея»). Для нас сейчас существенно, что цыплята не только бежали при предъявлении «ястреба», но также издавали крик «опасность с воздуха» (именно и только этот), и лишь в том случае, когда находились в группе — но не в одиночку. Следовательно, с помощью этого крика они не выражали собственные побуждения, а использовали сигнал именно для информирования других членов сообщества, позволяя им выбирать собственную стратегию поведения, которая будет зависеть от их личных «обстоятельств». Например, нехватка корма заставляет особь больше рисковать — она отреагирует на сигнал не так поспешно. Аналогичные результаты были получены и для приматов, в исследовании сигналов предупреждения об опасности у кольцехвостых, или кошачьих лемуров Lemur catta.
В других экспериментах у цыплят вырабатывали условную связь между нейтральным стимулом (красный свет) и появлением корма. После выработки условного рефлекса красный свет вызывал пищевой сигнал, свет иной длины волны — нет. Другой пример: у цыплят «система левого глаза» и «система правого глаза» выполняют различные функции. Первая отвечает в основном за пространственную локализацию удаленных объектов, вторая — за категоризацию объектов, например распознавание корма. Когда цыплятам проигрывали крик «опасность с воздуха», они пытались рассмотреть объект именно в верхней части поля зрения и именно при помощи левого глаза.
Существует множество доказательств тому, что сигналы-символы не зависят от контекста и соотносятся только с определенными категориями значимых событий внешнего мира животного. В книге Д. Чини и Р. Сифарта (Cheney D.L., Seyfarth R.M.. «How monkeys see the world: Inside the mind of another species», 1990) рассмотрен пример с мартышками верветками, чей «язык» сейчас активно исследуется. Верветки издают один и тот же «крик орла» и когда воздушный хищник так далеко, что может интерпретироваться лишь как «тревожный», но не «опасный» объект, и на самых последних стадиях его атаки, когда кричащие особи почти не имеют шансов спастись. Если бы крик маркировал определенный уровень опасности (связанный с появлением того, а не иного хищника), сигналы были бы резко различными.
Упомянутые выше кольцехвостые лемуры устойчиво реагируют криком «опасность с воздуха» на любое появление пернатых хищников и криком «наземная опасность» на появление хищных млекопитающих. Те и другие «обозначаются» своим специфическим сигналом независимо оттого, где относительно опасности находится сам лемур — на земле или на ветках, и оттого, как меняется скорость приближения хищника. Наконец, характер тревожного крика у цыплят зависит скорее от признаков потенциально опасного объекта, нежели от дистанции до этого объекта и, следовательно, оттого, как данная особь оценивает опасность для себя.
То же самое показали знаменитые опыты Р. Сифарта и Д. Чини с зелеными мартышками Cercopithecus aethiops. Адекватность и точность реагирования мартышек на сигнал об опасности определенного рода («орел», «змея», «леопард») полностью определяется тем, насколько соответствует акустическая структура данного крика некоторому «идеальному типу», который все особи этого вида распознают как сигнал с соответствующим «значением», а не просто «шум».
Именно сигналы-символы оказываются самым специализированным видом коммуникативных сигналов, вершиной эволюции сигнальных систем в конкретных филогенетических ветвях.
Так, у более примитивных, ранее сформировавшихся видов сурков (более длинные «ветви» филогенетического древа) есть единственный тревожный крик, который информирует лишь о том, что кричащая особь испытывает беспокойство, тогда как у более молодых видов появляются дифференцированные типы сигналов (рис. 7).
Рис. 7. Система сигналов, предупреждающих о различных опасностях, возникла у сурков в ходе эволюции. Филогенетическое древо рода Marmota построено по неповеденческим (молекулярным) признакам. Для каждого вида показан репертуар сигналов. Прямоугольники — различные типы сигналов: видно увеличение их числа при переходе от более древних видов к молодым. Изображение: «Химия и жизнь»

Еще раз подчеркнем, что по содержанию сообщений сигналы-символы противопоставляются сигналам-стимулам: первые информируют о ситуации извне, в которую втянуты особи, вторые о субъективной оценке напряженности ситуации самой особью. Сигналы-символы информируют партнера, а не воздействуют на него «здесь и сейчас». Сигналы-стимулы только воздействуют на реципиента, а не информируют его; причем сила и специфичность воздействия связана с уровнем мотивации и специфичностью мотивационного состояния особи, как и предполагают классические этологи (таблица 3). Наконец, действие первой категории сигналов связано с механизмом информирования, специфическим именно для сигналов, а второй — с механизмом стимуляции подобного подобным, которому подчиняются любые действия, обладающие некой выразительностью, а совсем не одни демонстрации.
Таблица. Сигналы-символы и сигналы-стимулы
1.Сигнал издается только на появление тех элементов мира, которые должны быть им «обозначены», и ни на что иное.
2.Система дифференцированных сигналов животного соответствует дифференцированным ситуациям, и это соответствие не нарушается, даже если меняются уровни общего возбуждения участников.

Подходящего русского аналога названий referential signals и motivational signals пока не существует. Ж. И. Резникова в своей книге («Интеллект и язык животных и человека: Основы когнитивной этологии», 2005) использует дословный перевод «категориальный сигнал». Нам кажется более подходящим по смыслу термин «сигналы-символы». В этом случае сохраняется то противопоставление сигналам-стимулам, которое существенно в любом исследовании роли сигналов как специализированных посредников в информационных обменах внутри сообщества.
К настоящему времени известно множество примеров коммуникативных взаимодействий, о которых можно уверенно сказать, что мы имеем дело с сигналами-символами, а не стимулами. Но это тема для отдельной статьи.
Что еще можно почитать о коммуникации животных:
1) Сайт Кристофера Эванса (C. S. Evans), где можно найти описания его работ по коммуникативным сигналам птиц.
2) The Animal Communication Project.
3) З. А. Зорина, И. И. Полетаева, Ж. И. Резникова. Основы этологии и генетики поведения. М.: изд.-во МГУ, Высшая школа, 2002.
4) Е. А. Гороховская.Этология: рождение научной дисциплины.
5) В. С. Фридман. Хронологическая таблица «История идей сравнительной этологии».
6) Marc D. Hauser. The Evolution of Communication. The MIT Press, 1997.